Paper
Organic & Biomolecular Chemistry
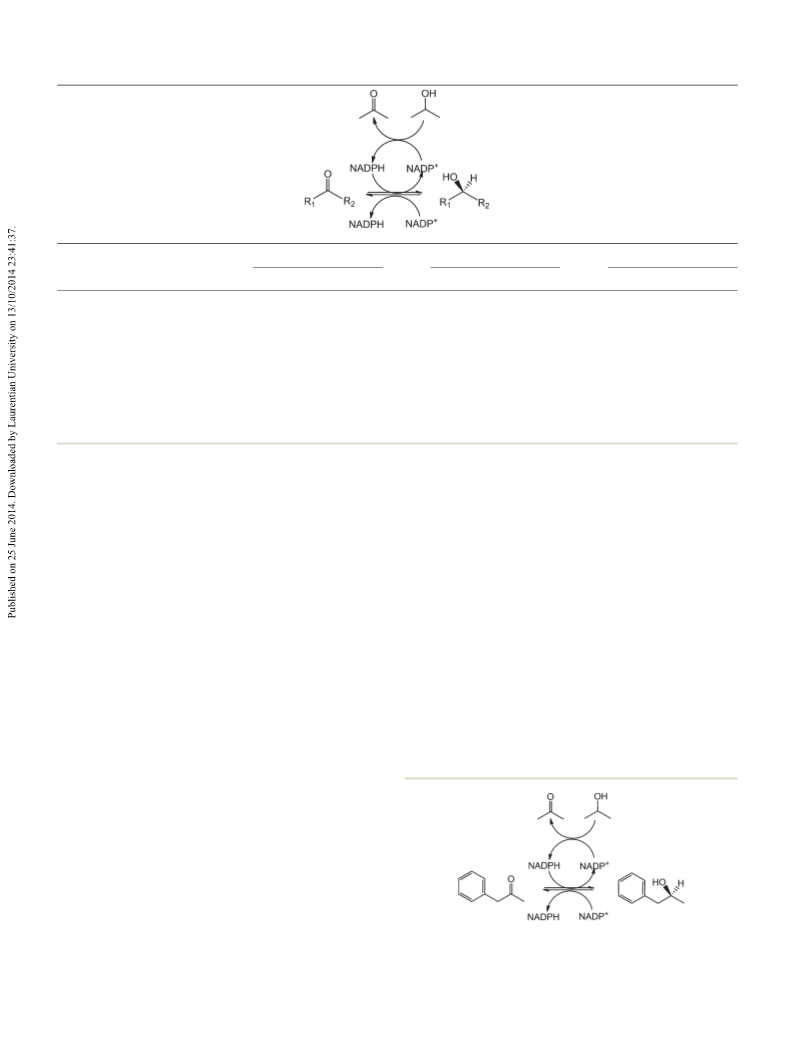
phenylacetone, 1-phenyl-2-butanone, and 4-phenyl-2-buta-
none, which represent remarkable improvement over the pre-
viously reported mutant, W110A TeSADH, which reduced
phenylacetone to (S)-1-phenyl-2-propanol in only 37% ee. One
of the new mutants, W110V TeSADH, shows extremely high E
and kcat/Km values of 134.5 and 45 300 M−1 s−1, respectively,
for (S)-1-phenyl-2-propanol. This new mutant TeSADH library
provides new insights in understanding the enzyme’s stereo-
specificity, and accompanied by further optimization of
TeSADH by mutations at other amino acid residues will make
this enzyme a more attractive catalyst for chiral aromatic
alcohol synthesis.
Materials and methods
General
Fig. 1 Crossed-eye stereo view of docking phenylacetone to W110V
TeSADH using Autodock Vina on the holoenzyme form of Thermoanaer-
obacter brockii ADH (PDB: 1YKF). The phenylacetone, shown in cyan, on
the left is the lowest energy pose for W110V SADH. The phenylacetone
molecule on the right side of the structure is the lowest energy pose for
wild-type TeSADH overlaid on the W110V structure. The gray sphere is
the catalytic zinc. The CPK stick structure is the NADP+.
Commercial grade solvents were used as obtained from the
suppliers. 1-Phenyl-2-butanone, 4-phenyl-2-butanone, and
NADP+ were used as purchased from commercial suppliers. All
buffer solutions were adjusted at room temperature. Capillary
GC measurements were performed on a GC equipped with a
flame ionization detector and either an HP chiral-20B column
(30 m, 0.32 mm [i.d.], 0.25 μm film thickness) or a Supelco
β-Dex 120 chiral column (30 m, 0.25 mm [i.d.], 0.25 μm film
thickness) using He as the carrier gas. Enzyme kinetic studies
were performed on a Varian Cary 1E UV-Vis spectrophotometer
equipped with a 6 × 6 Peltier temperature controlled cell block.
butanone, as well as their corresponding alcohols, with high
enantioselectivities and enantiospecificities, respectively. In
order to understand the molecular basis for these changes in
substrate specificity and stereochemistry, we conducted
docking studies for the mutants with phenylacetone and com-
Site-saturation mutagenesis
pared that with wild-type TeSADH. As
a representative
The codon NNK (N = GATC and K = GT) was used at W110 site
to get all the possible mutations using site saturation muta-
genesis based on the Quikchange (Stratagene) protocol. This
degenerate primer design approach gives 32 codons, and
theoretically gives access to all 20 amino acids in a single reac-
tion. Primer design was done using broad guidelines from Liu
and Naismith as well as Zheng et al.13,14 Partially overlapping
primer design was used in this case, and their guidelines
stated that there should be at least 8–18 overlapping bases, tar-
geted mutation in both forward and reverse primers, and at-
least one G or C should be at each of the termini.
example, phenylacetone was docked with the structure of
W110V Thermoanaerobium brockii SADH† created in silico
using Autodock Vina, and we found that the lowest energy
pose of phenylacetone docks within the active site, close to
residue 110, and the carbonyl oxygen is about 5 Å away from
the catalytic Zn2+, as shown in Fig. 1 (molecule in cyan on the
left side of the figure); however, for wild-type enzyme, phenyl-
acetone is unable to enter the active site, probably because the
large size of Trp-110 does not allow it to go in, and it is
blocked in the mouth of the active site (Fig. 1, molecule in
cyan on the right side of the figure). This is consistent with the
results obtained in this report, as well as previous ones,7,8 as it
is known that phenylacetone is not a substrate for wild-type
TeSADH, whereas it is a substrate for the other W110 TeSADH
mutants described herein.
Primers used: GGCAGGCN
GATGGTGTTTTTG
GAAAATTTMN̲ N̲ G̲ CCTGCCAGCATTCCACCGGAGTGCTGGTG.
̲N̲K̲AAATTTTCGAATGTAAAA-
The PCR reaction was done in a 50 μL tube containing 1 μL
of template (2–10 ng), 1 μM of primer pair, 200 μM of dNTP
and 3 units of Pfu Turbo DNA polymerase. The PCR cycle was
initiated by incubation at 95 °C for 5 min to denature the tem-
plate DNA, followed by 16 amplification cycles, each of 95 °C
for 1 min, 52 °C for 1 min, and 68 °C for 24 min, followed by
incubation at 68 °C for 1 h. The PCR reaction was then
digested by addition of 1 μL of Dpn I restriction enzyme
(10 U μL−1) with incubation for 1 h at 37 °C to digest the entire
parental methylated supercoiled DNA. After going onto the
further steps, we realized that we were having very high back-
ground of wild-type mutants, and that was attributed to incom-
Conclusions
In this paper, we used site-saturation mutagenesis to create a
library of mutants of TeSADH at residue W110, and we were
able to successfully obtain six very interesting mutants. Five of
them gave >99.9% ee in the enantioselective reductions of
†The amino acid sequences of SADH from T. ethanolicus and T. brockii are
identical.
5908 | Org. Biomol. Chem., 2014, 12, 5905–5910
This journal is © The Royal Society of Chemistry 2014

 Patel, Jay M.
Patel, Jay M.