Bilirubin photo-isomers
467
Although hepatic UGT1A1 activity is relatively low at
birth and shortly thereafter, the enzyme is not totally
deficient and its activity increases during the first *10
postnatal days [3, 4]. Whether photo-isomers of BR are
substrates for UGT1A1 or other hepatic enzymes is
unknown and has been difficult to determine because of
their instability and unavailability, as well as confusion in
the literature regarding their structures, preparation, and
identification. However, as will be shown, their instability
can be turned to advantage in the identification of their
metabolites. This paper investigates the glucuronidation
and phase II metabolism of BR photo-isomers in the rat. It
outlines simple methods for preparing BR photo-isomers in
sufficient purity and quantity for in vivo studies and
reappraises earlier structural assignments and methods for
the preparation of ‘‘purified’’ photo-isomers and BR
O
O
COOH
H
O
O
N
H
N
N
H
N
H
Fig. 3 Spirolactone structure assigned to photobilirubin II and
cyclobilirubin [23, 26]
compound. In addition to their relevance to neonatal
jaundice and phototherapy, the studies provide an expla-
nation for the formation of BR diglucuronide in normal
metabolism and question the structures and importance of
various hydroxylated bilirubins [18–21] proposed as
products of BR metabolism and phototherapy.
2
photoproducts. The studies show that the principal BR
photo-isomers are substrates for a uridine glucuronosyl
transferase (UGT)1 enzyme in vivo but, in striking contrast
to the parent (4Z,15Z)-isomer, undergo highly regioselec-
tive monoglucuronidation, with no diglucuronidation. The
stereoselectivity of glucuronidation seems to be dictated by
the ability to form intramolecular hydrogen bonds, as
demonstrated by studies on a simple stable model
Results and discussion
For metabolism studies, solutions of pure individual photo-
isomers or, at least, solutions highly enriched in the content
of specific isomers are essential. In preliminary work the
methods of Stoll and colleagues for preparing ‘‘purified’’
photo-isomers of BR-IXa were examined, beginning with
photobilirubins IA and IB, which had been assigned
structures (4Z,15E)-BR and (4E,15Z)-BR, respectively
2
The nomenclature and identification of BR photoisomers in the
literature is confusing. The photoisomer structures shown in Figs. 1
and 2 were established unambiguously by NMR and other spectro-
scopic and chemical methods published in 1982 and, for simplicity,
the trivial name ‘‘lumirubin’’ (CAN 83664-21-5 and 83729-98-0) was
assigned to structural isomers with the novel cycloheptadienyl ring
[
13], followed by photobilirubin II, originally identified as
a separable mixture of two stable conformational isomers
of (4E,15E)-BR but later assigned other structures,
including that of (Z)-lumirubin [23].
(
Fig. 2) [7, 8]. Previously the term ‘‘photobilirubin’’ had been used to
describe early photoproducts of BR [22], but that term became
redundant once the individual photoisomer chemical structures had
been established. Yet the term was not completely abandoned,
appearing in the literature as photobilirubins IA and IB, photoprod-
ucts isolated by Stoll et al. who thought them to be (4E,15Z) and
Photobilirubins IA and IB
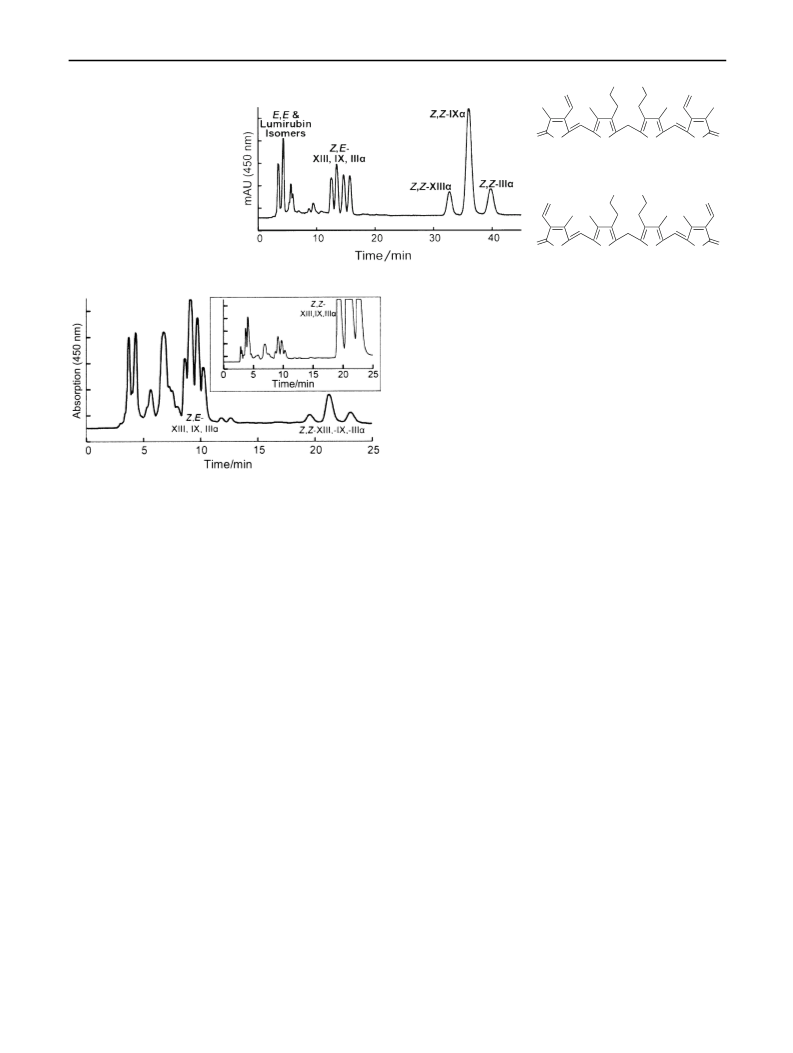
Samples were prepared by irradiation of BR in CHCl with
3
(
4Z,15E)-BR, respectively [13, 14]. However, as shown in this paper
a mercury lamp [13]. The final solution after irradiation
and the crude product obtained after extraction of photo-
isomers into acetone were dark green, indicating sub-
stantial by-product formation. Reversed-phase high-
performance liquid chromatography (HPLC) of the acetone
extract (Fig. 4) showed a complex mixture and revealed
that partial metathesis of the starting material had occurred.
Photobilirubin IA, isolated by thin-layer chromatography
those structure assignments are incorrect. Another photoproduct,
photobilirubin II, initially thought to contain two stable atropisomers
of (4E,15E)-BR [13, 14] was subsequently assigned either the
lumirubin structure (Fig. 2) or spiro-lactone structure (Fig. 3) [23],
and finally the lumirubin structure [24]. The lumirubin structure in
Fig. 2 is sometimes called (E,Z)-cyclobilirubin. Cyclobilirubin,
originally called ‘‘unknown pigment’’, was initially assigned the
(
[
4E,15E)-bilirubin structure [25], then the spiro-lactone structure
26], and eventually, in 1984 [27], the lumirubin structure elucidated
earlier [7]. The name is confusing because it implies that a (Z,Z)-
cyclobilirubin isomer could exist; however, such a structure is
stereochemically impossible. In 1987, Bonnett and Ioannou published
a table of structure/name correlations for the photoproducts isolated
by different investigators [28]. Unfortunately, there are errors in that
table. Adding further confusion, structures for several photoisomers
and for bilirubin glucuronides depicted in a more recent review [29]
are incorrect. The current paper uses unambiguous chemical nomen-
clature for configurational isomers and the trivial name lumirubin for
isomers with a cycloheptadienyl ring system linking two adjacent
pyrrolic rings formed by intramolecular cyclization of an endo vinyl
group. There is no longer a need for the ambiguous and confusing
photobilirubin or cyclobilirubin nomenclature.
(
(
TLC) as described [13], which had been assigned the
4Z,15E) structure, was found by HPLC to be a mixture
containing none of that isomer and with the approximate
bilirubin composition (based on relative peak areas) of 8 %
(
4E,15Z)-BR-XIIIa, 10 % (4E,15Z)-BR-IXa, 23 %
4Z,15Z)-BR-XIIIa, 47 % (4Z,15Z)-BR-IXa, and 11 %
(
(4Z,15Z)-BR-IIIa. Photobilirubin IB, which had been
assigned the (4E,15Z)-BR-IXa structure [13], contained
very small amounts of both (Z,E)- and (E,Z)-isomers of BR
and had the approximate composition (by HPLC) of 3 %
123

 McDonagh, Antony F.
McDonagh, Antony F.