Glucuronidation of Estrogen Stereoisomers
591
changes the spatial relations between 17-OH and the methyl group on Authorship Contributions
C18. It was not easy to find a good substrate for testing the latter
suggestion, and the reason 13-epiestradiol was added to the current
study was because it was the only possible currently available estrogen
in which the spatial organization of the 17-OH and 18-CH3 is
considerably different than in other estrogens. Nevertheless, even 13-
epiestradiol does not provide a very good test compound because the
conformational change in C13 leads to larger changes in the structure
of the entire estrogen molecule (Fig. 6).
Participated in research design: Sneitz, Finel.
Conducted experiments: Sneitz, Mosorin, Vahermo.
Contributed new reagents or analytic tools: Poirier, Vahermo.
Performed data analysis: Sneitz, Laakkonen, Vahermo.
Wrote or contributed to the writing of the manuscript: Sneitz, Finel.
References
Ayan D, Roy J, Maltais R, and Poirier D (2011) Impact of estradiol structural modifications (18-
methyl and/or 17-hydroxy inversion of configuration) on the in vitro and in vivo estrogenic
activity. J Steroid Biochem Mol Biol 127:324–330.
Barre L, Fournel-Gigleux S, Finel M, Netter P, Magdalou J, and Ouzzine M (2007) Substrate
specificity of the human UDP-glucuronosyltransferase UGT2B4 and UGT2B7. Identification
of a critical aromatic amino acid residue at position 33. FEBS J 274:1256–1264.
Cohen SL and Marrian GF (1936) The isolation and identification of a combined form of oestriol
in human pregnancy urine. Biochem J 30:57–65.
While the a configuration of C17 strongly increases the affinity of
UGT2B7 for estrogens, in the case of UGT1A10 it lowers the
enzyme’s affinity for estrogens, as far as can be judged from the K
m
values (Table 2). The configuration of the estriols’ C17 and/or C16
also affects the activity of UGT2B15 in estrogen glucuronidation,
except that the rates exhibited by this enzyme are relatively low and
the effects are more complex. When C17 of estradiol (17a-estradiol,
Itäaho et al., 2008, as well as enantio-estradiol, Sneitz et al., 2011) or
estriol (17-epiestriol, our study) is in the a configuration, UGT2B15
conjugates the 3-OH, the only member of the UGTs subfamily
Court MH, Zhang X, Ding X, Yee KK, Hesse LM, and Finel M (2012) Quantitative distribution
of mRNAs encoding the 19 human UDP-glucuronosyltransferase enzymes in 26 adult and 3
fetal tissues. Xenobiotica 42:266–277.
Eliassen AH, Spiegelman D, Xu X, Keefer LK, Veenstra TD, Barbieri RL, Willett WC, Han-
kinson SE, and Ziegler RG (2012) Urinary estrogens and estrogen metabolites and subsequent
risk of breast cancer among premenopausal women. Cancer Res 72:696–706.
Eliassen AH, Ziegler RG, Rosner B, Veenstra TD, Roman JM, Xu X, and Hankinson SE (2009)
Reproducibility of fifteen urinary estrogens and estrogen metabolites over a 2- to 3-year period
in premenopausal women. Cancer Epidemiol Biomarkers Prev 18:2860–2868.
Gall WE, Zawada G, Mojarrabi B, Tephly TR, Green MD, Coffman BL, Mackenzie PI,
and Radominska-Pandya A (1999) Differential glucuronidation of bile acids, androgens and
estrogens by human UGT1A3 and 2B7. J Steroid Biochem Mol Biol 70:101–108.
Höglund C, Sneitz N, Radominska-Pandya A, Laakonen L, and Finel M (2011) Phenylalanine 93
of the human UGT1A10 plays a major role in the interactions of the enzyme with estrogens.
Steroids 76:1465–1473.
Itäaho K, Mackenzie PI, Ikushiro S, Miners JO, and Finel M (2008) The configuration of the 17-
hydroxy group variably influences the glucuronidation of beta-estradiol and epiestradiol by
human UDP-glucuronosyltransferases. Drug Metab Dispos 36:2307–2315.
Katzenellenbogen BS (1996) Estrogen receptors: bioactivities and interactions with cell signaling
pathways. Biol Reprod 54:287–293.
2
B to do so, or at least this was our conclusion until testing the
glucuronidation of 13-epiestradiol (Fig. 7). In the case of UGT1A3, the
configuration change in C16 of estriol from a to b stimulates
conjugation but at the 16-OH, not at the 3-OH as it does when the
substrate is 17-epiestriol (Fig. 3B), estradiol, or 13-epiestradiol (Fig. 7).
UGT2B17, like UGT2B15, exhibited low rates in the glucuronida-
tion of the different estriols but an interesting stereoselectivity. As with
estradiols (Itäaho et al., 2008), UGT2B17 only conjugates the 17-OH of
estrogens when it is located “above” the plane of ring D. UGT2B17
failed completely to glucuronidate 17-epiestriol, the variant in which
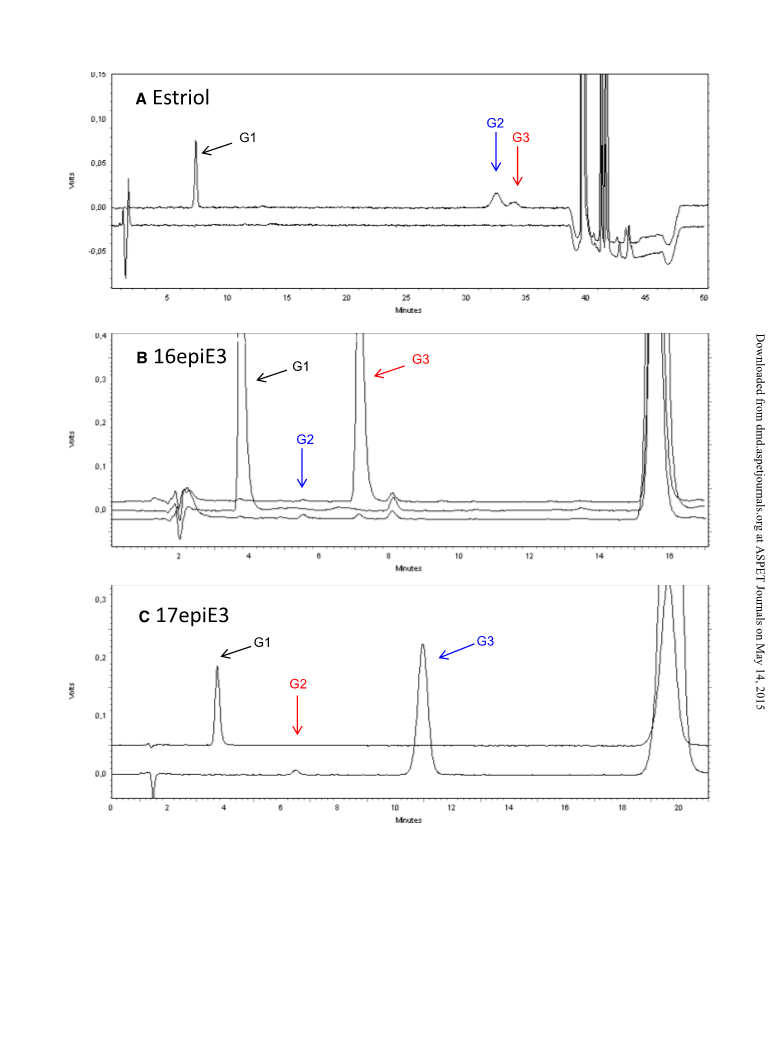
both C17 and C16 are in the a configuration (Fig. 1). In the case of 16-
epiestriol, where both the C16 and C17 are in the b configuration,
UGT2B17 conjugated both of them at similar rates (Fig. 3).
King CD, Rios GR, Green MD, and Tephly TR (2000) UDP-glucuronosyltransferases. Curr Drug
Metab 1:143–161.
Kurkela M, García-Horsman JA, Luukkanen L, Mörsky S, Taskinen J, Baumann M, Kostiainen
R, Hirvonen J, and Finel M (2003) Expression and characterization of recombinant human
UDP-glucuronosyltransferases (UGTs). UGT1A9 is more resistant to detergent inhibition than
other UGTs and was purified as an active dimeric enzyme. J Biol Chem 278:3536–3544.
Kurkela M, Patana AS, Mackenzie PI, Court MH, Tate CG, Hirvonen J, Goldman A, and Finel M
(
2007) Interactions with other human UDP-glucuronosyltransferases attenuate the con-
Aside from the glucuronidation site, the results also show that the
glucuronidation rate of the different estriols depends on the configu-
ration of C16 and/or C17, even if the conjugation target is the 3-OH, as
in UGT1A10 (Fig. 3). In this respect, it is interesting that a point
mutation in UGT1A10, F93G, largely changes the relation between
the structure of the estriol and its glucuronidation rate. The findings
with mutant 1A10-F93G also suggest that the differences in the
glucuronidation rate of the three estriols could not be assigned to
a small difference in the physicochemical properties among them.
The configuration change at C13 of estradiol changes the spatial
sequences of the Y485D mutation on the activity and substrate affinity of UGT1A6. Phar-
macogenet Genomics 17:115–126.
Lappano R, Rosano C, De Marco P, De Francesco EM, Pezzi V, and Maggiolini M (2010) Estriol
acts as a GPR30 antagonist in estrogen receptor-negative breast cancer cells. Mol Cell Endo-
crinol 320:162–170.
Lépine J, Bernard O, Plante M, Têtu B, Pelletier G, Labrie F, Bélanger A, and Guillemette C
(2004) Specificity and regioselectivity of the conjugation of estradiol, estrone, and their cat-
echolestrogen and methoxyestrogen metabolites by human uridine diphospho-
glucuronosyltransferases expressed in endometrium. J Clin Endocrinol Metab 89:5222–5232.
Longcope C (1984) Estriol production and metabolism in normal women. J Steroid Biochem 20
(
4B):959–962.
Melamed M, Castaño E, Notides AC, and Sasson S (1997) Molecular and kinetic basis for the
mixed agonist/antagonist activity of estriol. Mol Endocrinol 11:1868–1878.
Mukherjee TK, Nathan L, Dinh H, Reddy ST, and Chaudhuri G (2003) 17-epiestriol, an estrogen
metabolite, is more potent than estradiol in inhibiting vascular cell adhesion molecule 1
3
organization of the 17-OH and 18CH without changing the location
(
VCAM-1) mRNA expression. J Biol Chem 278:11746–11752.
of the 17-OH with respect to the plane of ring D (Fig. 6). The
glucuronidation results with estradiol and 13-epiestradiol suggest that,
with the exception of UGT2B17, the large change in the estradiol
molecule upon the configuration change at C13 does not affect the site
at which the different UGTs conjugate estradiol, but it does change the
reaction rate. The results with UGT2B17 were, however, surprising
because it also conjugated the 3-OH on ring A, an observation that has
never been reported before for UGT2B17 with any estrogen.
The results of this study shed new light on estrogen glucuronidation
by the human UGTs, particularly the structural components within the
estrogen substrate that, often differently, affect their glucuronidation
by individual UGT enzymes. These findings are expected to assist in
predicting the glucuronidation of drugs that resemble estrogens or
carry estrogen-like structural elements, such as the phase I metabolite
of exemestane (Sun et al., 2010). In addition, they will contribute to
a better understanding of the complex structure of the substrate-binding
site of the human UGTs and identification of the residues and other
structural properties that allow them to glucuronidate many different
substrates while retaining a considerable degree of specificity.
Ohno and Nakajin (2009) Determination of mRNA expression of human UDP-
S
S
glucuronosyltransferases and application for localization in various human tissues by real-
time reverse transcriptase-polymerase chain reaction. Drug Metab Dispos 37:32–40.
Radominska-Pandya A, Czernik PJ, Little JM, Battaglia E, and Mackenzie PI (1999) Structural
and functional studies of UDP-glucuronosyltransferases. Drug Metab Rev 31:817–899.
Ruggiero RJ and Likis FE (2002) Estrogen: physiology, pharmacology, and formulations for
replacement therapy. J Midwifery Womens Health 47:130–138.
Sneitz N, Court MH, Zhang X, Laajanen K, Yee KK, Dalton P, Ding X, and Finel M (2009) Human
UDP-glucuronosyltransferase UGT2A2: cDNA construction, expression, and functional charac-
terization in comparison with UGT2A1 and UGT2A3. Pharmacogenet Genomics 19:923–934.
Sneitz N, Krishnan K, Covey DF, and Finel M (2011) Glucuronidation of the steroid enantiomers
ent-17b-estradiol, ent-androsterone and ent-etiocholanolone by the human UDP-glucur-
onosyltransferases. J Steroid Biochem Mol Biol 127:282–288.
Starlard-Davenport A, Xiong Y, Bratton S, Gallus-Zawada A, Finel M, and Radominska-Pandya
A (2007) Phenylalanine(90) and phenylalanine(93) are crucial amino acids within the estrogen
binding site of the human UDP-glucuronosyltransferase 1A10. Steroids 72:85–94.
Sun D, Chen G, Dellinger RW, Sharma AK, and Lazarus P (2010) Characterization of 17-
dihydroexemestane glucuronidation: potential role of the UGT2B17 deletion in exemestane
pharmacogenetics. Pharmacogenet Genomics 20:575–585.
Tukey RH and Strassburg CP (2000) Human UDP-glucuronosyltransferases: metabolism, ex-
pression, and disease. Annu Rev Pharmacol Toxicol 40:581–616.

 Sneitz, Nina
Sneitz, Nina