Morakinyo et al.
417
Scheme I.
tem of enzymes and the other based on the flavin-containing
monooxygenase (8–10). In the presence of these enzyme
systems and cofactor-like compounds such as sulfide and
hydroxylamine, cysteamine is oxidized to the sulfinic acid
derivative, hypotaurine. Most metabolic pathways, however,
give hypotaurine as a precursor to taurine (the sulfonic acid).
Scheme I encompasses nearly all possible reactions of
cysteamine in the physiological environment.
Cysteamine and its major metabolite, hypotaurine, are
known to be excellent scavengers of HOCl and other reac-
tive oxygen species (ROS) and are more likely to act as anti-
oxidants in vivo than taurine (3, 11, 12). These ROS can
cause damage to DNA, proteins, and lipids in addition to
causing radical damage and oxidative stress in animal cells
(11, 13, 14). In addition to protection against radical damage
in DNA, cysteamine can also act as a repair agent for DNA
through the formation of the protective RSSR•–, which then
reacts with the DNA•+ radical ion to regenerate DNA and
form cystamine, the cysteamine disulfide (2, 3). The ability
of cystamine to reversibly form disulfide links with the
sulfhydryl groups at or near the active sites of enzymes is
also important in regulation of several essential metabolic
pathways (15, 16).
[R1] H2O2 + Br– + Myeloperoxidase/H+ ÷
HOBr + H2O
Taurine and its precursors, e.g., hypotaurine and
cysteamine have been suggested as possible moderators of
HOBr toxicity by forming longer-lived and less oxidizing
products in their reactions with HOBr (24).
Experimental
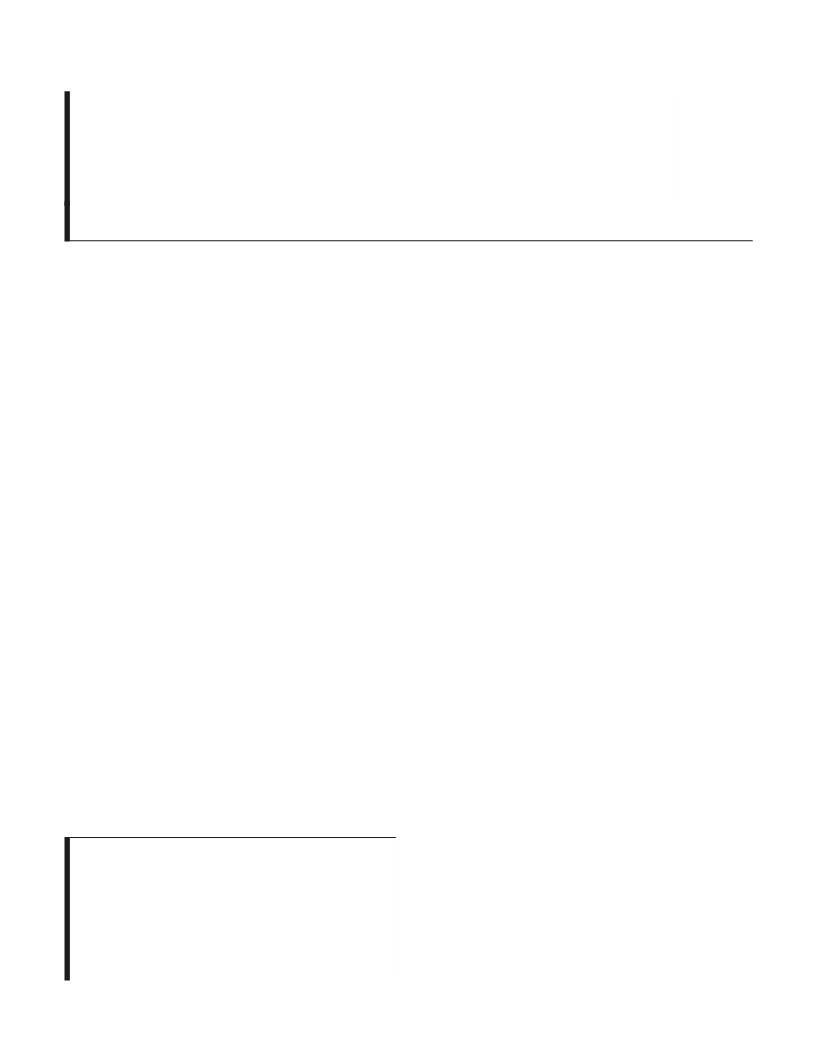
Our research group embarked on a series of studies to elu-
cidate the kinetics and mechanisms of the oxidation of
cysteamine and its S-oxides. Previous studies done in our
laboratory on the cysteamine sulfinic acid, hypotaurine, have
shown that it reacts quite rapidly with chlorite to give a mix-
ture of taurine, monocloro- and dichloro-taurine (17). Oxida-
tion is believed to occur simultaneously at the sulfur center
(to give the sulfonic acid) and at the nitrogen center (to give
the chloramines). The oxidation of taurine by chlorite (18)
and bromate (19) showed that oxidation occurs only on the
nitrogen centre to give the corresponding N-derivatives. The
reaction was exceedingly slow, even with the most powerful
oxidizing agents such as acidic bromate. The formation of
N-bromo- and N-chloro-taurines is suggested as a possible
mechanism by which taurine can moderate the oxidative tox-
icity of halogens and hypobromous and hypochlorous acid in
the slightly basic physiological environments (20).
A recent study from our laboratory on the oxidation of
cysteamine by iodine and acidic iodate showed that these
mild oxidants only oxidized cysteamine to its dimer,
cystamine (21). In this study, stronger oxidants bromine and
acidic bromate have been used. The aim of the study re-
ported in this manuscript was to investigate whether these
oxidants also produced cystamine as a viable product or
whether the oxidation proceeded to hypotaurine or all the
way to taurine as well as the N-bromotaurines. Bromate is a
precursor to HOBr, a particularly damaging oxidizing spe-
cies in the physiological environment. Stimulated granulo-
cytes produce oxidizing agents (especially H2O2) and secrete
granular proteins into the extracellular medium, which con-
tributes to their antimicrobial, cytotoxic, and cytolytic activi-
ties (22). Each group of cells contains a specific peroxidase,
which catalyzes the reactions of hydrogen peroxide with
halogens. The enzyme, myeloperoxidase, which is abundant
in neutrophils, catalyzes the oxidation of Br– ions by H2O2
to yield HOBr as a reactive and oxidatively damaging prod-
uct (23).
Materials
Cysteamine hydrochloride (CA, 2-aminoethanethiol hydro-
chloride) 98%, cystamine, bromine, potassium iodide, potas-
sium bromide (Sigma-Aldrich), sodium perchlorate (98%)
(Acros), sodium bromate, perchloric acid (72%), soluble
starch, sodium thiosulfate, and hydrochloric acid (Fisher)
were used without further purification. The concentration of
bromine was determined by standardization against
thiosulfate solution after addition of excess acidic iodide.
Spectrophotometry was used as a complementary technique
by measuring bromine absorbance at 390 nm where the ex-
tinction coefficient had been deduced to be 142 mol–1 L cm–1.
This standardization was carried out before each series of ki-
netic experiments owing to the volatile nature of bromine. CA
solutions were prepared just before use and not kept for more
than 24 h. All solutions were prepared using distilled and
deionized water from a Barnstead Sybron Corporation water-
purification unit. Inductively coupled plasma mass spectrome-
try (ICPMS) analysis showed negligible concentrations of
iron, copper, and silver and approximately 1.5 ppb of cad-
mium and 0.43 ppb in lead as the most abundant metal ions,
which was not enough to affect the overall reaction kinetics
and mechanism (25).
Methods
All experiments were carried out at 25.0 0.1 °C and a con-
stant ionic strength of 1.0 mol/L (NaClO4). CA, sodium per-
chlorate, and perchloric acid solutions were mixed in one
vessel and bromate (or bromine) solutions in another. A
PerkinElmer Lambda 25 UV–vis spectrophotometer interfaced
to a Pentium IV computer was used for most of the kinetics
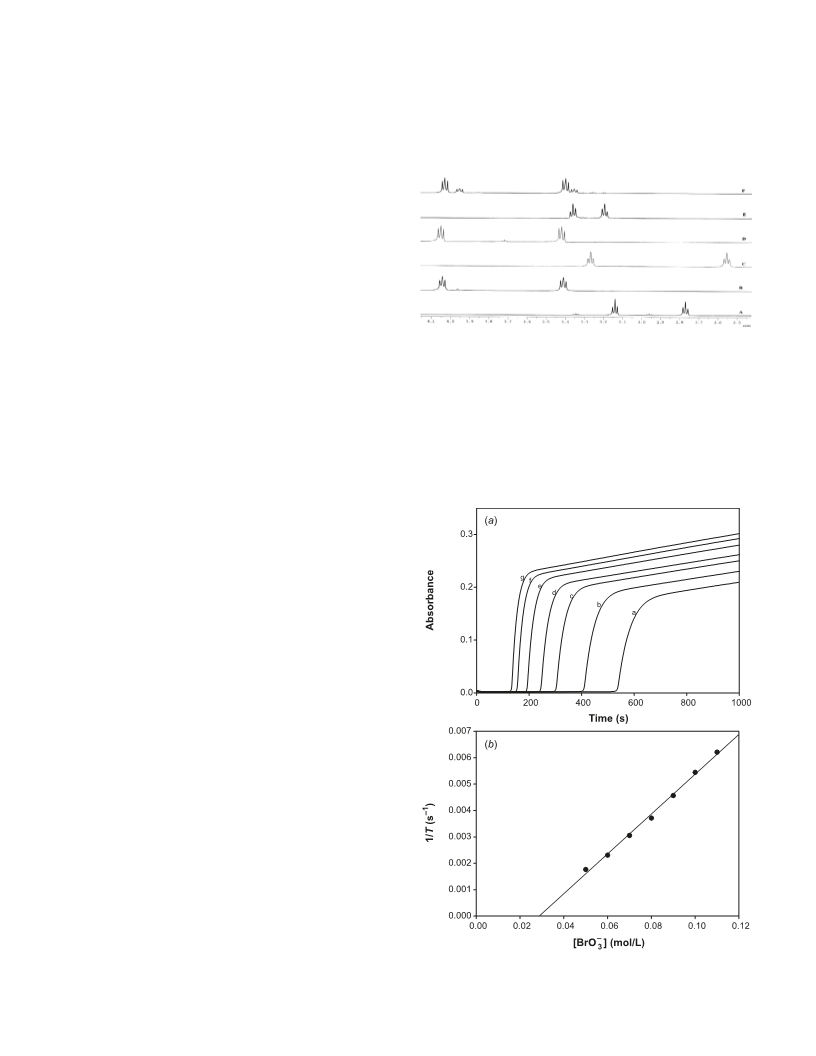
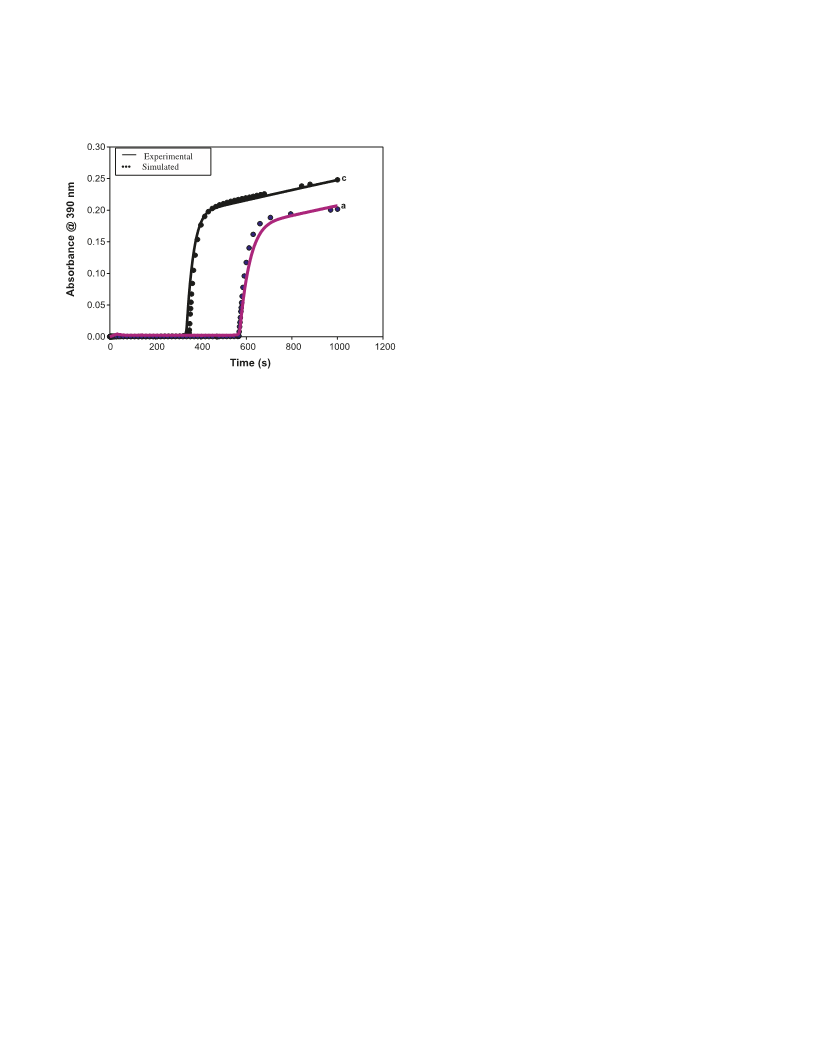
–
and stoichiometric determinations. The slower BrO3 – CA re-
actions were monitored on the UV–vis spectrophotometer by
monitoring bromine absorbance at 390 nm. Faster reactions
were followed on a Hi-Tech Scientific SF61 – DX2 double
mixing stopped-flow spectrophotometer.
© 2008 NRC Canada

 Morakinyo, Moshood K.
Morakinyo, Moshood K.